La sintesi moderna (speciale #DarwinDay)
Fu il 1859, quando negli scaffali delle librerie di tutta l'Inghilterra, venne pubblicata un'opera che generò inizialmente scalpore, ma in seguito fu l'artefice di una portentosa rivoluzione. Stiamo parlando di "Of Origins of Species" e del suo autore chiamato Charles Darwin, che ha aperto le "porte" studiando e osservando quel fenomeno che oggi è ancora attivo e plasma gli organismi viventi chiamato "evoluzione".

Buongiorno a tutti, qui Doc e benvenuti nel mio speciale #DarwinDay dove parliamo della Sintesi Moderna.
Prima di cominciare è d'uopo effettuare una piccola premessa, dell'opera di Darwin del XIX secolo, solo alcuni aspetti sono tutt'ora utilizzati, altri no.
Ho dovuto aggiungere questa postilla perché esistono ancora degli individui che sono ancora convinti che nel mondo accademico si utilizzi ancora l'opera di Darwin così come sia.
I PRIMI PASSI VERSO LA SINTESI MODERNA
Siamo sul finire del XIX secolo, oltre al modello evolutivo descritto da Darwin, c'era anche il modello di Lamarck, erano in 2 a contendersi il posto finché un giorno un tale August Weismann (colui che intuì la meiosi che avviene nelle cellule sessuali e non solo, dopo 15 anni dalla sua intuizione, James Sutton vedrà questo processo), confutò a livello sperimentale il concetto di "ereditarietà" di Lamarck.
 |
| August Weismann (1834-1914) |
Lamarck sosteneva che le modificazioni che un organismo vivente accumula durante tutta la sua vita, si trasmettano alla prole, cosa non dimostrata.
Anche nell'opera di Alfred Russell Wallace c'è un accenno al fatto che le modificazioni accumulate durante l'intera vita non si trasmettano alla prole.
D'altro canto, Darwin proponeva il "mescolamento" per spiegare il motivo della somiglianza tra i discendenti e gli antenati, ovvero che le caratteristiche in una popolazione si vadano a mescolare fra loro.
Comunque il modello di Lamarck aveva dalla sua l'ortogenesi (da Willhelm Haacke, ovvero lo sviluppo evolutivo in modo lineare di un carattere o di un organo, contiene un'elevata percentuale di finalismo), oltre al saltazionismo di Hugo de Vries.
Verso gli inizi del XX secolo, il mondo accademico era in tumulto perché erano state riscoperte delle leggi di un monaco d'istanza in un monastero di Brno (città presente nell'attuale Repubblica Ceca) e che nel 1864, aveva già provato ad esporle, ma il mondo scientifico non riusciva a seguirlo ed era disinteressato, si trattava di Gregor Mendel.
Mendel fece degli esperimenti non solo sulle piante dei piselli, ma anche sulle api e altri animali.
Il XX secolo, sarà il testimone della nascita della genetica.
Nel 1910, ci fu Thomas Hunt Morgan con l'osservazione delle mutazioni del colore degli ocelli dei moscerini della frutta (Drosophila melanogaster) e osservò a livello sperimentale che l'ereditarietà dei caratteri nei cromosomi che pubblicò come lavoro nel 1915.
Nel 1918, ci fu Ronald Fisher che pubblicò un lavoro statistico sull'eredità mendeliana.
Nel 1937, abbiamo un lavoro "Genetics and the Origin of Species" da parte di Theodor Dobzhansky, uno degli allievi di Morgan dove abbiamo l'unione tra il modello di Darwin e le leggi di Mendel.
LA SINTESI MODERNA VERA E PROPRIA
Verso gli anni '30 e '40 del XX secolo, si viene ad affermare la "Sintesi Moderna" che conosciamo noi oggi e che è utilizzata tutt'ora livello accademico.
Sviluppata grazie ai lavori di grandi personalità come John Maynard Smith, George C Williams, Ernst Mayer (di lui parliamo dopo, perché ha dato ulteriori contributi).
La Sintesi Moderna contiene:
- La selezione naturale di Charles Darwin
- Le 3 leggi dell'ereditarietà dei caratteri di George Mendel
- La genetica delle popolazioni con modelli matematici
- L'osservazione dei fossili in paleontologia
- La teoria degli equilibri punteggiati di Eldredge e Gould
- I grafici e modelli di Matoo Kimura
- L'EVO-DEVO
Il "Neodarwinismo" è il movimento culturale nato con August Weismann che ha portato alla Sintesi Moderna che è un modello scientifico con cui descriviamo il fenomeno dell'evoluzione.
Affermare che siano la stessa cosa, è commettere la fallacia logica dell'argomento fantoccio.
La Sintesi Moderna presenta 3 principi:
- La variabilità genetica individuale è frutto delle mutazioni che sono il "motore dell'evoluzione"
- L'evoluzione è un fenomeno che interessa LA POPOLAZIONE e opera NON SU UN SINGOLO INDIVIDUO, ma sull'INTERO PATRIMONIO GENETICO. L'insieme di tutti gli alleli (ovvero delle forme varianti di un solo gene) presenti in una popolazione costituisce il pool genico della popolazione.
- La selezione naturale preserva mutazioni vantaggiose, i cui portatori di codeste mutazioni aumenteranno in frequenza di generazione in generazione ed elimina più o meno velocemente quelle svantaggiose.
I CONTRIBUTI DI ERNST MAYR
Nel 1991, Ernst Mayr pubblicò un libro intitolato "One Long Argument" dove suddivise il lavoro di Darwin in 5 punti che adesso vado ad elencare:
- CAMBIAMENTO CONTINUO
- DISCENDENZA COMUNE
- MOLTIPLICAZIONE DELLE SPECIE
- GRADUALISMO
- SELEZIONE NATURALE
CAMBIAMENTO CONTINUO
Il mondo vivente non è "statico" come lo pensava il fissismo biologico, ma in continuo cambiamento, ovvero le caratteristiche degli organismi viventi variano di generazione in generazione.
Le testimonianze di codesti cambiamenti, sono stati osservati a livello morfologico nei vari fossili rinvenuti e inoltre i suddetti ci forniscono anche testimonianze di cambiamenti ambientali.
Purtroppo la fossilizzazione è un evento raro perché la carcassa dell'organismo deceduto, deve contenere elementi fossilizzabili come gusci, denti, ossa e parti dure.
Codesta carcassa deve essere seppellita velocemente da detriti, oppure da uno strato di argilla, in modo da essere sottratta all'azione degli agenti chimici, fisici e biologici.
Non abbiamo trovato molti fossili proprio per questo motivo (perché la fossilizzazione è rara), oppure alcuni devono ancora essere studiati in maniera più approfondita, altri li abbiamo persi a causa dell'erosione, perché sono fuoriusciti in un periodo in cui non c'era l'essere umano oppure in un periodo in cui la paleontologia non era ancora nata e non sono stati raccolti o venivano studiati, ma con il concetto dell'evoluzione.
L'evoluzione si basa di più sullo studio dei cambiamenti, anche nei viventi, ma in larga parte sui fossili.
DISCENDENZA COMUNE
Darwin arrivò ad intuire che tutte le specie animali e vegetali discendessero da un antenato comune e concepì l'idea dell'albero filogenetico.
L'albero con le sue ramificazioni rappresentava come si sviluppa e si diversifica la vita sul nostro pianeta.
Questo albero non è tutto bello e rigoglioso, ha delle parti dove le fronde mancano perché i rami da dove esse si sviluppano sono secchi, ovvero privi di vita. Quei rami rappresentano le specie che ormai si sono estinte.
Ora introduciamo il concetto di OMOLOGIA.
 |
| Esempio di omologia |
L'omologia è quando un organo ha la stessa origine embrionale però è presente in organismi differenti con forme differenti e funzioni differenti.
Nell'immagine qui sopra potete osservare un esempio di omologia ovvero la differenza tra i vari arti in diversi animali.
Queste strutture omologhe rappresentano caratteristiche che sono ereditate, con qualche modifica, provenienti da strutture appartenenti ad antenati comuni.
Poi esiste un secondo concetto l'ANALOGIA.
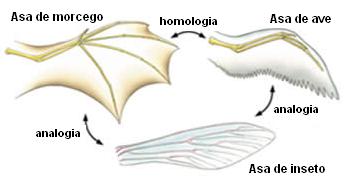 |
| Esempio di analogia |
L'analogia è quando le strutture svolgono la stessa funzione come l'ala di un pipistrello, di un dittero e di un uccello, ovvero volare.
Però codeste strutture possiedono origini embrionali diverse.
Poi c'è il contributo dell'EVO-DEVO che trovate in un altro mio post cioè qui.
L'omologia è collegata alla FILOGENESI o se preferite lo "studio che ricostruisce i rapporti tra i taxa viventi ed estinti".
Come facciamo
Per partire con la filogenesi, bisogna utilizzare la variabilità dei caratteri tra i membri del gruppo stesso, per ogni carattere bisogna stabilire quale variante del suddetto fosse presente nell'antenato comune più recente di tutto l'intero gruppo.
Questa forma del carattere la chiamano "stato ancestrale" e tutte le altre varianti, succedute per evoluzione le chiamano "stati derivati" del carattere.
Per capire (ricordo che è un lavoraccio), quale carattere sia antico o nuovo si fa attraverso dei confronti con quello che si chiama "outgroup" che è un gruppo simile dal punto filogenetico al gruppo che si studia che è "l'ingroup".
Se un carattere è condiviso sia dall'ingroup e sia dall'outgroup, allora per l'ingroup è ancestrale.
Se nessuno stato del carattere è condiviso sia nell'ingroup e sia nell'outgroup allora è antico per l'outgroup .
Vengono usati anche diversi outgroup e se un carattere è assente in tutti gli outgroup per il confronto, sono caratteri derivati.
La filogenesi comprende termini come "clado", dove esso comprende una linea ancentrale e tutte le specie discendono da codesta linea ancestrale.
Oggi utilizziamo quella che viene chiamata FILOGENESI MOLECOLARE, si basa sul concetto che le sequenze nucleotidiche del DNA, siano ereditate e con esse anche le sequenze che porteranno alla sintesi di catene amminoacidiche e in seguito alle proteine.
Più recente è la separazione tra 2 specie e più simili saranno le sequenze nucleotidiche e amminoacidiche.
Queste sequenze necessarie per i confronti sono disponibili in apposite banche dati internazionali.
Si tiene conto di 2 tipi di DNA, ovvero quello mitocondriale, di derivazione materna che presenta un tasso di mutazione più elevato di quello presente nel nucleo della cellula eucariote.
Con quello possiamo ricostruire rapporti filogenetici con specie (devono essere affini) che si diversificate di recente.
A volte vengono usati i "loci microsatelliti" ovvero quelle sequenze di DNA che sono ripetute e non codificanti.
Il secondo tipo di DNA è quella sequenza che codifica per l'RNA ribosomiale che presenta un tasso di mutazione più lento, con quello ricostruiamo i rapporti tra specie molto antiche e quando si sono diversificate, dell'ordine di milioni di anni.
Si utilizzano anche i geni regolatori aventi sigla HOX, essi regolano lo sviluppo embrionale.
MOLTIPLICAZIONE DELLE SPECIE
Quando compare un punto di ramificazione nell'albero filogenetico, vuol dire che una specie ancestrale si è divisa in specie diverse.
La specie, intesa nella sua definizione classica, presenta alcuni requisiti:
- La discendenza comune
- La riproduzione deve essere compatibile
- Deve avere una coesione genotipica e fenotipica all'interno.
In natura, 2 specie diverse non possono incrociarsi perché esistono delle barriere che possono essere pre o post zigotiche.
Le pre-zigotiche impediscono la fecondazione o ostacolano l'accoppiamento, quelle post-zigotiche impediscono che lo zigote ibrido diventi un adulto e pure fertile.
Ciò che impedisce l'accoppiamento è:
- ISOLAMENTO DELL'HABITAT: 2 specie che occupano differenti habitat all'interno della stessa specie possono possono incontrarsi raramente o non incontrarsi affatto. Questa situazione si verifica anche se le 2 specie non risultano isolate da barriere fisiche (esempio: una montagna).
- ISOLAMENTO TEMPORALE: le specie si accoppiano durante ore diverse del giorno, in differenti stagioni o in anni diversi, non c'è l'incontro tra i loro gameti.
- ISOLAMENTO COMPORTAMENTALE: i rituali di corteggiamento e altri comportamenti esibiti da una specie possono essere vere e proprie barriere riproduttive, anche se sono strettamente imparentate.
Ciò che ostacola la fecondazione è:
- ISOLAMENTO MECCANICO: alcune differenze anatomiche possono impedire il successo riproduttivo di un eventuale accoppiamento.
- ISOLAMENTO GAMETICO: gli spermatozoi sono incapaci di fecondare la cellula uovo di una specie diversa.
- RIDOTTA SOPRAVVIVENZA DEGLI IBRIDI: i geni delle differenti specie parentali possono interagire in modo da alterare lo sviluppo o diminuire la sopravvivenza degli ibridi.
- RIDOTTA FERTILITÀ DEGLI IBRIDI: gli ibridi pur mostrando un ottimo grado di vitalità possono essere sterili. Se i cromosomi delle 2 specie parentali differiscono per numero o struttura, negli ibridi la meiosi non è in grado di produrre gameti. Dal momento che gli ibridi sterili non sono in grado di concepire neppure in caso di accoppiamento con una delle specie parentali, non può esserci il flusso genico da una specie all'altra.
- SCOMPARSA DEGLI IBRIDI: in alcuni casi gli ibridi della prima generazione sono vitali e fertili, ma quando si accoppiano fra loro o con membri delle specie parentali la prole delle generazioni successive, tende a essere sterile e debole.
Queste barriere fra le popolazioni evolvono gradualmente, quindi è necessario che le popolazioni debbano rimanere isolate fisicamente per un lungo intervallo di tempo.
Però prima che si instaurino del tutto le "barriere riproduttive" una popolazione in fase di diversificazione può ritornare in contatto e quindi incrociarsi e avviene il rimescolamento genico.
Il processo che porta alla formazione di nuove specie prende il nome di "Speciazione" e ne esistono di 3 tipi.
La prima è la SPECIAZIONE ALLOPATRICA, è quando 2 popolazioni vivono l'un l'altra in aree geografiche separate, però sono composte da individui dalla stessa specie, incapaci accoppiarsi, ma possono farlo una volta rimosse le barriere geografiche.
2 popolazioni separate, possono evolversi indipendentemente e adattarsi ai nuovi ambienti, però nascono barriere riproduttive oltre a nuovi genotipi e fenotipi.
Quando la barriera viene rimossa le 2 specie non possono più riprodursi, avviene in 2 modi:
- SPECIAZIONE PER VICARIANZA: scatenata da un cambiamento climatico o un evento geologico, l'habitat dove vive una specie si frammenta e vengono create barriere impenetrabili che separano popolazioni diverse dal punto di vista geografico. A volte un singolo evento vicariante può frammentare simultaneamente più specie diverse e nonostante la frammentazione la popolazione rimane integra.
- IL PRINCIPIO DEL FONDATORE: sono coinvolti sono pochi individui di una specie, essi si disperdono e raggiungono nuovi territori e distanti, in questi nuovi luoghi non esistono altri membri della loro specie e creano una nuova popolazione. All'inizio ha dimensioni molto limitate che causa un cambiamento veloce e drastico della struttura genetica della popolazione ancestrale e del fenotipico.
Il secondo tipo è la SPECIAZIONE SIMPATRICA, dove l'origine di una nuova specie avviene senza isolamento geografico.
Perché avvenga ci deve essere una riduzione del flusso genico da particolari fattori:
- POLIPLOIDIA: una nuova specie si genera da un evento accidentale anomalo verificatosi nel corso di una mitosi, il corredo cromosomico risulta soprannumerario.
- SELEZIONE SESSUALE: si ipotizza la presenza di sottogruppi che si specializzano nell'utilizzo di particolari risorse, per questo motivo si divergono geneticamente dando a luogo a nuove specie.
- DIFFERENZIAMENTO DELL'HABITAT: si verifica quando una sottopopolazione presenta una mutazione che la induce a utilizzare in maniera diversa l'habitat o le risorse rispetto alla popolazione parentale.
Esiste un terzo tipo di speciazione: la speciazione PARAPRATICA, intermedia tra le prime 2, è quando si ha una locazione allopatrica, ma c'è un contatto in modo che una delle 2 specie supera la barriera con successo.
Nella speciazione esiste un fenomeno chiamato RADIAZIONE ADATTIVA, dove si formano nuove specie ecologicamente differenziate, come ad esempio i mammiferi che è iniziata nell'era mesozoica ovvero nel Giurassico ed ha accelerato dopo l'estinzione dei dinosauri non-aviani e i fringuelli nelle isole Galapagos.
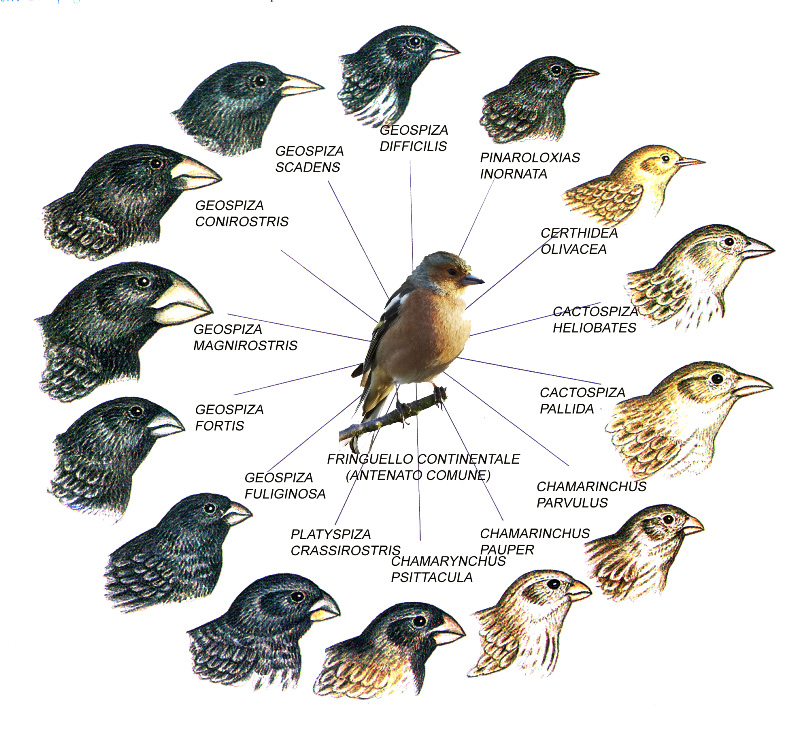 |
| Radiazione adattiva dei fringuelli delle Galapagos |
GRADUALISMO
Piccole differenze, man mano che si accumulano, alla lunga portano alla comparsa di una nuova specie.
Mayr distingue 2 tipi di gradualismo ovvero quello di popolazione e fenotipico.
- GRADUALISMO DI POPOLAZIONE: in una popolazione si affermano nuovi caratteri grazie a un aumento della loro frequenza da una frazione limitata, fino alla maggioranza della popolazione stessa. Abbiamo molte prove di questo gradualismo.
- GRADUALISMO FENOTIPICO: afferma che i nuovi caratteri, anche quelli che portano alle divergenze maggiori con i fenotipi ancestrali, sono prodotti una serie di piccoli passi graduali, accumulati in centinaia di migliaia di generazioni. Qui ci sono ancora discussioni in atto.
Darwin nella sua opera, parlava di "gradualismo filetico" dove ci sono una sequela di forme intermedie che collegano fenotipi ancestrali a quelli odierni, però questo tipo di gradualismo è raro da trovarlo nei fossili.
Per tentare di spiegare queste "mancanze" nel 1972, Niles Eldredge e Stephen Jay Gould formularono la "teoria dell'equilibrio punteggiato", con questo modello, i cambiamenti fenotipici non sono costanti, nel senso che a volte si concentrano in tempi brevi, altre volte dopo lunghi periodi di stasi.
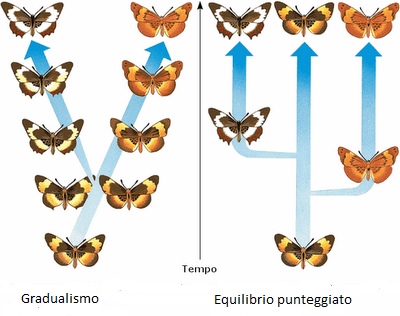 |
| La differenza tra i vari gradualismo |
SELEZIONE NATURALE
Il processo cardine del modello di Darwin, con questo concetto si da una spiegazione naturale all'origine e all'adattamento di tutte le caratteristiche anatomiche, fisiologiche e comportamentali e dello sviluppo che rendono gli organismi più adatti a sfruttare le risorse ambientali per sopravvivere e riprodursi.
Darwin le ha sviluppate da 5 osservazioni e 3 deduzioni, alcune delle osservazioni e deduzioni sono state influenzate dall'opera di un economista britannico chiamato Thomas Robert Malthus, che scrisse un saggio nel 1798 delle popolazioni.
Secondo Malthus, le popolazioni di piante e animali (anche quelle umane) aumentano in un numero che va oltre la capacità di sopportazione del sistema.
Le 3 osservazioni sono
- Gli organismi hanno un ampio potenziale riproduttivo che permette una crescita esponenziale della popolazione.
- Le popolazioni naturali di solito non crescono, ma si mantengono pressoché costanti nel tempo.
- Le risorse naturali sono limitate.
Queste 3 osservazioni convergono nella deduzione che i membri di una popolazione c'è una continua lotta per l'esistenza.
Le altre 2 osservazioni:
- In una popolazione tutti gli individui mostrano variazioni
- Le variazioni sono ereditabili, i discendenti tendevano ad assomigliare ai genitori, Darwin non capiva le ragioni di questo fatto
Queste convergono in altre 2 deduzioni, ovvero che gli individui diversi mostrano sopravvivenza e capacità riproduttiva differenziate che favoriscono i caratteri vantaggiosi.
L'ultima deduzione è che la selezione naturale, agendo su molte generazioni, gradualmente produce nuovi adattamenti e nuove specie.
Come esempio esiste il caso della Biston betularia, una falena che esiste in 2 forme, chiara e scura.
Normalmente la forma chiara si mimetizza meglio sulla corteccia che è chiara e quella scura viene predata maggiormente.
Durante la seconda rivoluzione industriale in Inghilterra, le sorti si sono invertite, il pulviscolo derivante dall'utilizzo del carbone ricoprì la corteccia e la falena forma chiara questa volta è lei ad essere maggiormente predata e si salva quella scura.
Quando poi il carbone cessò di essere usato, la corteccia ritornò ad essere chiara e la forma scura ritornò ad essere predata come prima.
 |
| Le 2 forme della falena |
LA MATEMATICA DELL'EVOLUZIONE
Parafrasando un noto fisico, che diceva che nell'evoluzione non c'è la matematica, peccato per lui che ci sia eccome, per esempio abbiamo l'equazione dell'equilibrio di Hardy e Weinberg.
L'equazione di Hardy e Weimberg sostiene che in assenza di forze perturbatrici, ovvero in condizioni di equilibrio, la frequenza di ciascun allele di una popolazione si trasmettono inalterate da generazione in generazione, secondo codesta equazione purché ci sia evoluzione non ci devono essere condizioni di equilibrio.
Le "forze perturbatrici" sono:
- Mutazioni
- Flusso genico
- Dimensioni piccole
- Accoppiamenti non casuali
- Selezione naturale
Esistono poi le equazioni di George Robert Price, genetista delle popolazioni, sviluppate nel 1970, descrive i cambiamenti della selezione naturale o artificiale e anche i cambiamenti dovuti al flusso genico.
Poi ci sono i modelli di sostituzione delle basi azotate di Matoo Kimura che studiano il meccanismo delle mutazioni genetiche.
Bene l'articolo finisce qui e noi ci vediamo alla prossima.
